MTHFR677T and 1298C Play Independent but Opposite Effects in Stroke - Additive Co-Effect of two Intragenic Alleles
Introduction
Methylenetetrahydrofolate reductase (MTHFR) is essential for homocysteine metabolism [1] and elevated total homocysteine is associated with a variety of cardiovascular diseases (CVD) including stroke [2-9] although such associations may not be causal [9]. Two variants in the coding region of MTHFR gene, 677C>T (p. Ala222Val) and 1298A>C (p. Glu429Ala), are prevalent in North American, European and Asian populations, and the 677T is associated with elevations in total homocysteine levels [7-8,10-13]. For this reason, the 677T variant has been extensively investigated for association with CVD, including stroke, by using a variety of different populations [14-24], and including several meta-analyses [5,7-8, 25-27]. The association between T variant (especially homozygote of TT) and CVD, including stroke, was described in a large number of these studies [28-29] whereas, no association was seen in others, including a large meta-analysis [5,30-37]. Compared with the 677T variant, the 1298C variant, located 2.1 kb from the 677T position, has not been extensively studied, and most studies published so far show no association with neither elevated homocysteine levels [13,38-39] nor CVD [10,18,24] and IS [7,40]. The relationship between the 677T and 1298C variants remains a puzzle and continues to be the subject of much debate.
Etiologies of multifactorial disease usually involve multiple genetic events, environmental factors and the combination of both. For a single genetic variant with a strong effect, it is usually easy to demonstrate association with a related disease, and such association can easily be replicated in the different ethnic populations. However, a moderate or mild genetic effect modified by other genetic or environmental factors could present complicated results in association studies involving different ethnic populations due to differences in genetic background and environmental living conditions. A recent meta-analysis revealed a model of gene - environment interaction in which the 677T effect on homocysteine levels was highly dependent on folate status [8]. The discordant results from previous association studies with 677T indicates that the effect of the 677T variant could be enhanced/diminished by other prevalent genetic change(s) acting either in concert or in opposition.
In the present study, the 677T and 1298C variants were genotyped in 1429 ischemic stroke (IS) patients and 1171 healthy age and sex-matched controls from the North Han Chinese population. Considering the potential for interaction between the two intragenic variants, our study mainly focused on the association between IS and all genotypic combinations among the four different alleles of 677C/T and 1298A/C. The combined genotype analysis enabled us to determine the independent genetic effects of 677T and 1298C variants in the circumstance of non-compound status, as well as a possible co-effect of the two variants in a compound status.
Methods
Studied Population:
1,429 consecutives IS patients were recruited from 18 hospitals within Henan province during the period from February 2006 to March 2007. Clinical diagnosis of IS patients was based on the WHO criteria for IS (1998) plus evidence from MRI or CT exam suggestive of ischemic lesions corresponding to the neurological deficits. The 1,171 normal controls were selected from ethnically and geographically matched individuals without history of myocardial infarction or stroke who presented in hospitals for routine health examinations. Written informed consent was obtained from all patients. Ethics approval for this present study was granted by Committee of Ethics Boards of Zhengzhou University.
Genotyping of 677C>T (rs1801133) and 1298A>C (rs1801131 ):
Genomic DNA was isolated from peripheral blood using standard salt precipitation methods [41]. Genotyping of 677C>T and 1298A>C were conducted using the Taq Man SNP genotyping technology on real-time PCR (ABI Prism® 7000 Sequence Detection System) which was described in our previous studies [42-43]. The primers and probes for 667>T and were obtained from the validated TaqMan SNP genotyping kit supplied by Applied Biosystems (ABI; Foster City, CA).
Prevalence determination and Statistical Analysis:
The prevalence of each gene variant was calculated by counting the total carrier frequency including heterozygotes and homozygotes. Independent effect of each variant was determined by calculating the sum of prevalence for all possible combined genotypes. The allele frequencies were determined by gene counting. Tests of Hardy-Weinberg equilibrium (HWE) were carried out for all loci among Stroke patients and controls separately by the Chi-square test. Risk ratios (RR) were calculated as a measure of the relative risk for Stroke and were given with 95% confidence intervals. All statistical analyses were performed by using R 3.4.4 (R Core Team, Vienna Austria). Linkage disequilibrium between the 677C>T and 1298A>C variants was calculated as D’, which ranges from 0 (no linkage disequilibrium) to 1 or -1 (complete linkage disequilibrium).
Results
General Genotyping Analysis
The 677C/T and 1298A/C variants were genotyped in the entire studied population including the 1492 IS patients and 1171 age and sex matched healthy controls from the North Han Chinese population. The genotype prevalence and allele frequency of each of 677C/T and 1298A/C variants were analyzed individually. Details about the prevalence distributions of all possible genotypes from the two variants and the allele frequencies of the variants in patients and controls are given in Table 1. The 677T variant presents the major allele with allele frequency of 0.5948 in the North Han Chinese population (control population). Without considering the fact of possible co-existence with 1298C variant allele, the 677T variant was associated with IS by comparing the allele frequency (RR=1.0865, 95% CI: 1.0407 1.1343, P=0.0002), and prevalence analysis of the sum of all genotypes with T allele (CT + TT) (RR=1.0668, 95% CI: 1.0324-1.1024) P=0.0001) between patients and controls. With respect to individual genotypes of 677C/T, the association was only observed for TT (RR=1.1308, 95% CI: 1.0259-1.2464, P= 0.0133) but not for CT. This suggests that the T allele predisposes for IS, but the susceptibility becomes possible only with homozygous status (TT). Dosage effect of T allele was also suggested by comparing with the risk ratios (RR) generated from CT and TT analysis (1.0155 vs 1.1308) in patients and controls. On the contrary, the 1298C allele has a relatively low frequency (0.1337) in the North Han Chinese population (control population). The C allele, in general, was negatively associated with IS by comparing the allele frequency (RR=0.8456, 95% CI: 0.7311-0.9781, P=0.0239) and the prevalence of all genotypes with C alleles (AC + CC) (RR=0.8448, 95% CI: 0.7333-0.9733, P=0.0195) between the patients and controls. This suggests a protective effect of C allele against IS. Regarding each individual 1298A/C genotype, the negative association was only achieved in the genotype of AC (RR=0.8438, 95% CI: 0.7271-0.9793, P=0.0254). The differences CC genotype frequency failed to achieve statistical significance, possibly due to the low prevalence of CC genotype in both patients and controls (0.0161% vs 0.0188%).
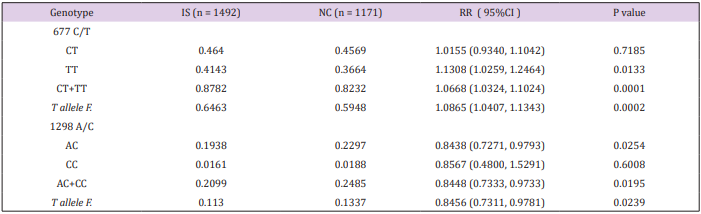
Table 1: Combined Regressions by [K/(Ca+Mg); CaMgNPK]. Causes of death with group numbers of Statistics Finland.
Combined Genotype Analysis
The prevalence of combined genotypes among the each of the four alleles from 677C/T and 1298A/C variants was investigated in patients and controls, and the results are depicted in Table 2. The combined genotypes of 677CC/1298AA, representing the homozygous wild type of both variants, was significantly higher in controls compared with IS patients. This supports the wild type status of both 677C and 1298A variants in the Chinese population. The combined genotypes, 677TT/1298AA and 677CC/1298CC denotes homozygous variants in one allele and homozygous wild type in the other allele. These two combined genotypes present ideal situations to determine independent effects of 677T and 1298C variants, respectively. The 677TT/1298AA genotype, but not the 677CT/1298AA genotype is significantly associated with IS (RR=1.3619, 95% CI: 1.2242-1.5151, P=1.35E-08), which suggests that an independent effect of the T allele predispose to IS but the susceptibility only presents in the TT homozygous status (dosage effect). In term of the 1298C allele, the 1298AC/677CC genotype is negatively associated with IS (RR=0.6845, 95% CI: 0.5043-0.9291, P= 0.0150), and suggests an independent protective effect for IS.
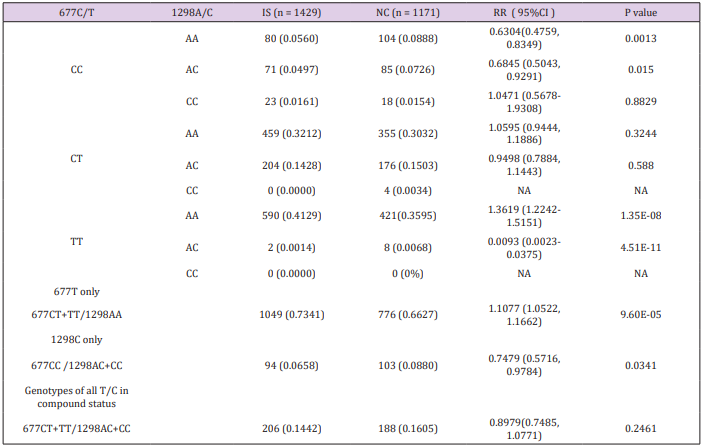
Table 2: Regression by [K/(Ca+Mg);CaMgNPK] explained highly significantly (p< 0.001) CVD (91%), Land traffic accidents (87 %), Non-CVD (86 %), Digestive diseases (excluded alcohol related diseases) (81 %), Diseases of the respiratory system (81 %), Diabetes (62 %), Suicides (62 %) and Assault (41 %).
The protective role of 1298C is effective in the heterozygote status. Unfortunately, examination of 677CC/1298CC genotype failed to demonstrate a negative association with IS, which may be due to the low prevalence of this genotype in patients (0.0161) and controls (0.0154). Although the 677TT/1298AC genotype was associated with significant negative risk for IS (RR=0.0093, 95% CI: 0.0023-0.0375, P=4.51E-11), given the small number of subjects with this genotype caution must be exercised in the interpretation of this finding in the absence of confirmation. Moreover, there was no 677TT/1298CC nor 677CT/1298CC genotypes in the studied population. It was therefore not possible to examine for a co-effect involving 677T and 1298C at this step of the analysis.
Grouped Combing Genotypes Analysis
A strategy of grouped combined genotype analysis was used to confirm the independent effects of 677T and 1298C and investigate for a possible co-effect of both variants. The results of the association study with each grouped combined genotype are given in Table 2. In this study, the grouped genotypes 677CT+TT/1298AA represent all genotypes in which the 677T is not in compound with 1298C. Similarly, the grouped combined genotypes of 677CC/1298AC+CC represent all genotypes where the 1298C is not in compound with 677T. Finally, the grouped combined genotypes of 677CT+TT/1298AC+CC represents all genotypes where 677C and 1298T coexist (double homozygote, double heterozygote and homozygote heterozygote combination). Significant distribution disequilibrium of 677CT+TT/1298AA (RR=1.1077, 95%CI: 1.0522- 1.1662, P=9.60E-05) and 677CC/1298AC+CC (RR=0.7479, 95% CI: 0.5716-0.9784, P=0.034) were detected in patients compared with controls. This confirms an independent but functionally opposite effect of 677T and 1298C for IS. Examination of all genotypes where the 677T and 1298C alleles are found in compound status (677CT+TT/1298AC+CC) showed that both the risk effect of 677T and protective effect of 1298C were neutralized in patients compared with the controls when both variants are in compound status (RR 0.8979, 95% CI: 0.7485-1.0771, P=0.2461). This is evidence of a genetic additive co-effect acting in opposite directions with respect to disease risk.
Discussion
Genetic predisposition in multifactorial disease, such as IS, results from a complicated co-effect from multiple genetic events. The multigenic effect can be due to either an additive co-effect from multiple independent genes, or synergistic co-effects from genegene and/or allele-allele interactions [44-45]. Most of the genetic susceptibilities described so far play a moderate or weak role in disease pathogeneses and the effects of susceptibilities are easily influenced by other genetic modifiers. Additive co-effects is result from the sum of all individual effects from multiple functioning independent genes, and a synergistic co-effect comes from two or more functioning related gene-gene interactions (genetic modifying). To demonstrate a co-effect from two or more genes, a large study population is necessary in order to gain sufficient statistical power for combined genotype distribution analysis. Furthermore, a population with less genetic heterogeneity is also critical because of sharing the similar genetic modifier(s). In this study, the 677C/T and 1298A/C were genotyped in 1492 IS patients and 1171 age and sex matched healthy controls from the North Han Chinese population. To ensure less heterogeneity in genetics and environment, all samples were collected from Henan Province and the majority of individuals originated from the local population.
The contradictory results concerning 677T in previous association studies can be explained if there is co-existence of either genetic modifiers or another genetic event(s) with opposite effect. The susceptibility of 677T depends on prevalence of such coexistent genetic change(s) which could vary among different ethnic populations. The 677T and 1298C genotypes are both located in the coding region of the MTHFR gene. Considering a possible intragenic allelic interaction, the two variants, in this study, were analyzed together focusing on the different combined genotypes among the four different alleles between the 677C/T and 1298A/C. High allelic frequencies of 677T and 1298C and a relatively large sample size (2663) in the study gives sufficient statistical power to analyze the distribution of each combined genotype in patients and controls. Through an analysis of the combined genotyes where the two variants are not in compound status, we were able to determine the independent but opposite effects of 677T and 1298C for IS. The independent effects of 677T and 1298C were further confirmed by the grouped combined genotype study (677CT+TT/1298AA and 677CC/1298AC+CC). Furthermore, the opposite effects of 677T and 1298C were indirectly confirmed in the study with grouped combined genotype, 677CT+TT/1298AC+CC, which also suggests an additive co-effect from 677T and 1298C. We, therefore, have reason to believe that discordant results in previous association studies with MTHFR may be the result of variable allele frequencies of 677T and 1298C among the different studied populations. A continued effort to validate the findings from this study is under way.
Systemic inflammation has been reported as a major predisposing factor for CAD and stroke. Khalighi et al. [46] recently reported that the 677T and 1298C have opposite effects on systemic inflammation where the 677T was significantly associated with increased NLR level and the1298C showed the opposite effects and tended to direct lower levels of NLR and PLR than 1298A. Our result provided evidence from a different angle to support independent but opposite effects of 677T and 1298C. Further study is needed to investigate the relationship between NLR level and IS and determine if the risk effect of 677T and protective effect of 1298C for IS are mediated through a mechanism involving the pathogenesis of systemic inflammation.
Meta-analysis is a method that enables pooling data from smaller inconclusive studies to yield greater statistical power to allow one to quantify genetic risks more precisely. However, it is difficult for meta-analyses, in practice, to obtain data from all possible combined genotypes from the multiple studies based on the available published literatures. Moreover, variable gene frequencies among ethnically different populations could also be another challenge for meta-analyses to pin point a possible alleleallele/gene-gene interaction.
At present it is not possible to exclude 677T and 1298A being in linkage disequilibrium with other gene(s) associated with IS because these two variants are closely located in nucleotide position (same Linkage Disequilibrium block). However, such possibility is low considering the significantly higher prevalence of the combined genotype, 677CC/1298AA, in the control population (RR=0.6304; 95% CI: 0.4759-0.8349, P=0.0013). Furthermore, the independent protective effect of 1298C allele has been determined in this study. Although the sample size used in this study provides sufficient power to perform combined genotype analysis, a validation study with increased samples size is required and underway by our research group. The 677T was previously shown to affect folate metabolism and plasma homocysteine levels. The design of the present study did not permit a comparison of the genotypes with biochemical parameters. A study in this area to elucidate the relationship with biochemical risk factors would be of value in future research efforts.
Conclusion
The results of this study clearly demonstrate that
a) 677T and 1298C play independent but opposite effects in IS when the two variants are not in compound status.
b) The 677T independently predisposes for IS, but the IS susceptibility presents only in the homozygous status (dosage effect).
c) The 1298C is negatively associated with IS, and therefore plays a protective effect.
The two functionally opposite effects from the 677T and 1298C can be neutralizing when the two variants are in the compound status. This latter result also provides evidence of an additive coeffect for the two intragenic alleles.
For more Articles: https://biomedres01.blogspot.com/



No comments:
Post a Comment
Note: Only a member of this blog may post a comment.